Surfactant production by epiphytic bacteria, Lysinibacillus sphaericus BiBi1 and Proteus mirabilis strain BiBi8 and their potential for hydrocarbon degradation
Oso, Simisola Olabisi, Ashade, Ahmeed Olalekan, Fanu, Blessing Seide
Department of Microbiology , Lagos State University , Ojo, Lagos, Nigeria
In this study, the ability of Lysinibacillus sphaericus BiBi1 and Proteus mirabilis BiBi8 isolated from and identified using biochemical and 16S rRNA molecular typing from leaves of Celosia argentea, were tested for their ability to utilise diesel as sole carbon source and also produce surfactants. The growth dynamics of isolates on diesel over 13-day incubation on diesel mineral salt medium revealed that Strain BiBi1 grew exponentially from day 0 to Day 8, with growth reaching its peak at day 10. A gradual decrease in the pH of the medium was observed from day 0 to day 13. Proteus mirabilis had a steady growth rate on diesel from day 0 to day 10 where growth was at its peak on day 12. Bacterial strain BiBi1 and BiBi8 showed alpha and beta hemolysis on blood agar and also recorded E24 % value of 51.28 and 76.32 respectively. The surfactants produced were characterized using high performance liquid chromatography, Lysinibacillus sphaericus produced rhamnolipids and subtilis as its surface biomolecule whereas Lysinibacillus sphaericus, in addition to rhamnolipid also produced lipopeptides. The results obtained from this study shows that Lysinibacillus sphaericus strain BiBi1 and Proteus mirabilis strain BiBi8 have potential for remediation of hydrocarbon contaminated environments.
Key word: Biodegradation; diesel; hydrocarbon; Enrichment; Surfactants
Petroleum hydrocarbons are known to be composed of complex mixtures of alkanes, aromatics, asphaltenes, and resins (Abbasian et al., 2015). They pose a threat and great hazard to life in terrestrial and aquatic environments (Ezeonu et al, 2102, Uddin et al,2021). This is due to their persistent nature of remaining in the environment for a long period leading to an alteration in the physical and chemical properties of the soil and also, the microorganisms known to be indigenous to the soil (Abdel-Shafy and Mansour, 2016). Over the years, various methods such as chemical, physical, and thermal treatments have been employed to treat hydrocarbons. These methods have been proven to be expensive and also require secondary treatment of these hydrocarbons (Das and Chandran 2011; Bilal and Iqbal,2020). Due to these reasons, it is important to find methods that are environmentally friendly and cost-effective when treating these recalcitrant hydrocarbons.
An example is the use of microorganisms in biodegradation which converts petroleum hydrocarbons to CO2, H2O, and biomass which are less harmful to the environment (Das and Chandran, 2011; Bala et al, 2022). Biodegradation is a natural process that utilizes microorganisms to clean up environmental contaminants (Varjani and Upasani, 2017). During biodegradation, different microorganisms such as bacteria, fungi, protozoa, and algae utilize contaminants such as petroleum hydrocarbons as sources of carbon and energy. Bacteria are the commonly used ones and examples are Pseudomonas, Sphingomonas, Rhodococcus, Micrococcus, Alcaligens, Aeromicrobium, Flavobacterium, Bacillus and Williamsia (Oso et al., 2019, Imron et al, 2020). Enzymes produced by microorganisms play an important role in the biodegradation process by breaking down the waste material into safe environmental compounds (Karigar and Rao, 2011). However, for biodegradation to be effective factors such as temperature, nutrients, and pH should be optimal to allow for the growth of microorganisms (Karamalidis et al., 2010). The first step in the degradation of hydrocarbons by microorganisms involves bacterial membrane oxygenase which requires direct contact with the substrate (Das and Chandran 2011; Xu et al., 2018).
The bioavailability and concentration of the hydrocarbon and environmental factors are determinants of the successful degradation of hydrocarbons. The degradation of hydrocarbons using a mixed culture of microorganisms has also proven to be effective because they possess a pool of catabolic genes which are required for an accelerated rate of degradation of hydrocarbons that are not found in a pure culture. Therefore, a mixture of mixed cultures of microorganisms can lead to higher degradation rates of hydrocarbons (Janbandhu and Fulekar 2011, Abbasi et al., 2017; D’ugo et al., 2021).
The degradation of petroleum hydrocarbons is carried out by an enzyme system, attachment of microbial cells to the hydrocarbon, or by the production of surfactants (Das and Chandran 2011; Xu et al., 2018). The use of surfactants to enhance the degradation of hydrocarbons by increasing the solubility of hydrocarbons which in turn increases the bioavailability of the hydrocarbons to the microorganisms has proven to be an effective method of hydrocarbon degradation (Oso et al., 2019; Oso et al., 2021). Surfactants are amphiphilic compounds known to reduce surface tension by accumulating at the interface of immiscible fluids therefore increasing the surface area of insoluble compounds leading to increase bioavailability and enhance hydrocarbon degradation (Li and Chen, 2009; Burch et al., 2014).
The utilization of hydrocarbons by surfactant-producing bacteria as a degradation method however depends on the type of hydrocarbons, type of surfactants produced by the microorganisms, and the characteristics the microorganisms possess (Hua and Wang 2014; Sah et al., 2022). The phyllosphere, known to be the above-ground organs of plants is a habitat for a wide range of microorganisms such as bacteria, fungi, yeast, and oomycetes (Vorholt, 2012). Bacteria are the most abundant ranging from 10-4 – 10-5 bacteria per mm2. Examples of known bacteria on leaves are Pseudomonas, Rhodococcus, Acinetobacter, Bacillus, Methylobacterium, Pantoea, Sphingomonas, etc (Remus-Emsermann and Schlechter, 2018).
Microorganisms in the phyllosphere can produce surfactants that aid their effective survival and exploration of the phyllosphere e.g. syringafactin produced by Pseudomonas syringae (Krimm et al., 2005; Burch et al., 2014). Surfactants produced by these bacteria inhabiting the phyllosphere are useful in the areas of agriculture, cosmetics, pharmaceuticals, and the environment. These surfactants produced by bacteria in the phyllosphere aid the removal of organic compounds such as petroleum hydrocarbons by decreasing surface and interfacial tensions (Abdoli et al, 2025, Patel et al, 2021). The surfactant-producing bacteria in the phyllosphere possess hydrocarbon-degrading genes and are capable of degrading hydrocarbons (Xu et al 2018). Though, it is not unknown if these epiphytic bacteria gain fitness by utilising the hydrocarbons present on plant leaf surfaces (Vorholt, 2012).
These leaves in the phyllosphere are covered by a hydrophobic cuticle which protects against biotic and abiotic stresses such as water loss reduction and UV radiation (Riederer and Schreiber 2001, Krajšek et al., 2011, Zeisler and Schreiber 2016, Sivakumar et al., 2020). The cuticle present on leaves is known to be a composite structure of phenolics, mineral elements, and polysaccharides cutin and cuticular waxes. Cuticular waxes are a polymer containing crosslinked very long aliphatics and soluble aliphatics (Pollard et al. 2008, Yeats and Rose 2013). It also acts as the primary interface for microorganisms on leaves and insect interaction (Riederer & Schreiber, 2001; Yeats et al., 2012). Cuticles differ in composition, structure, and thickness depending on the type of plant (Ridge 1997; Barthlott et al., 1998) . A large percentage of these bacteria listed above inhabiting the leaf surface is known to be hydrocarbon degraders and might be able to utilise the waxes on the leaf surface as a carbon source. However, there is no evidence that these bacteria can utilize the waxes present on the leaf surface (Remus-Emsermann et al., 2014; Schlechter et al., 2019). . These bacteria also perform many ecological roles such as fighting against plant diseases and host-specific pathogens for trigonistic functions in the microbiota (Bonaterra et al., 2022). In this study, the ability of two bacterial strains, Lysinibacillus sphaericus strain BiBi1 and Proteus mirabilis strain BiBi8 isolated from Celosia argentea leaves to utilize hydrocarbon and produce surfactants were looked into extensively.
Study site and Sample collection
The sample Celosia argentea, used for the study were collected from Lagos State University (LASU) farmland behind the Faculty of Education. The site has witnessed continuous farming practice for over three decades. It has a coordinate (Latitude 6º28′29′′ N and Longitude 3°11′52′′ E). The sites was selected among different farmland on campus due to its biological richness, accessibility, and the prominence of Celosia argentea as a plant species.
Isolation of diesel degrading bacteria
Diesel degrading bacteria were isolated from the sample (Celosia argentea) via continuous enrichment technique by aseptically punching out one-hundred plugs from 20 leaves (Celosia argentea) with the aid of cock borer, the plugs were washed in a sterile 20 ml normal physiological saline. Carbon free mineral salt medium (MSM) was prepared as prepared as described by Habe et al., 2003. The MSM constituent l-1 were 0.22 g/l Na2HPO4, 0.8 g/l KH2PO4, 3.0 g/l NH4NO3, 0.2 g/l MgSO4·7H2O, 0.05 g/l FeCl3·6H2O, 0.01 g/l CaCl2·2H2O and 0.05 g/l yeast extract and pH of the medium adjusted to 7.2. Trace elements solution (1 ml/L) were filter sterilized using membrane filter 0.45 μm and added aseptically to the medium as described by Bauchop and Elsden (1960), fungal growth was suppressed using 50 μg/ml nystatin. Forty-five (45) ml MSM was transferred into replicate 250 ml Erlenmeyer flask supplemented with 1 ml sterile diesel serving as sole carbon source, and 4 ml aliquot containing the washed cell were added. Incubation were done in the dark and flask subjected to constant agitation in an orbital shaker at room temperature (27 ± 2.0°C) for 14 days until there was growth (turbidity). Following 3 consecutive transfers at shorter intervals (7 to 14 days), hydrocarbon degraders were isolated by plating out the final flask on Luria-Bertani (LB) agar. The colonies that appeared were further purified by subculturing repeatedly on LB agar.
Growth of Isolates on diesel
Sets of flasks containing 50 ml carbon free mineral salt medium supplemented with 0.5 ml diesel (1%) were prepared. The test organism was inoculated into the flasks and, incubated at ambient temperature (27 ± 2◦C) for 13-d under constant agitation in the shaker. The control flask containing mineral salt medium and diesel with heat killed cells. At intervals of 2 day flask were randomly selected and their cell density determined via optical density measurement (Beckman Coulter DU 720, Fullerton, CA) while entire content of flask sacrificed. The growth patterns of isolates were plotted using Prism version 5.0 (GraphPad Software, San Diego, CA).
Identification of Isolates
Bacterial isolates were identified using colony morphology, cellular and biochemical characteristics using the taxonomic scheme of Cowan and Steel’s (Barrow and Feltham, 1995). The biochemical test carried out include Gram staining, catalase, oxidase, sugar utilization, indole, motility and hydrogen silphide production. Biochemical characterization was complemented with 16 rRNA molecular typing.
Molecular identification of Isolates
Genomic DNA was extracted from the cultures on sterile broth using the Quick-DNATM Fungal/Bacterial Miniprep Kit (Zymo Research, Catalogue No. D6005). The 16S target region was amplified using One Taq® Quick-Load® 2X Master Mix (NEB, Catalogue No. M0486 with the primers 16S-27F forward AGAGTTTGATCMTGGCTCAG and 16S-1492R Reverse CGGTTACCTTGTTACGACTT. The PCR products were run on a gel and cleaned up enzymatically using the EXOSAP method. The extracted fragments were sequenced in the forward and reverse direction (Nimagen, BrilliantDyeTM Terminator Cycle Sequencing Kit V3.1, BRD3-100/1000) and purified (Zymo Research, ZR-96 DNA Sequencing Clean-up KitTM, Catalogue No. D4050). The purified fragments were analyzed on the ABI 3500xl Genetic Analyzer (Applied Biosystems, ThermoFisher Scientific) for each reaction for every sample. BioEdit Sequence Alignment Editor version 7.2.5 was used to analyze the. ab1 files generated by the ABI 3500XL Genetic Analyzer and results were obtained by a BLAST search (NCBI) (Stephen et al., 1997). The 16S rRNA gene sequences were then deposited in the GenBank Database (accession number CP026120; CP053718).
Screening for surface active biomolecules
Hemolysis test
The method described by Satpute et al., 2008 was used to assay for the hemolytic activity of the isolates. Bacterial isolates were streaked on Luria-Bertani (LB) agar supplemented with 5% fresh sheep blood and incubated at (27 ± 2 °C) for 48-72 h. Plates were observed for alpha, beta and gamma-hemolysis. Greenish clearing zones around the colonies indicated alpha-hemolysis, while complete clearing zone around the colonies indicated beta hemolysis. gamma-hemolysis was characterized by lack of changes around the colonies. Hemolysis was presumptive for surfactant production.
Emulsification index
The emulsification indices (E24) was measured using the method described by Ilori et al., 2005. surfactant activity is measured by adding 2ml of diesel to 2 ml of cell-free extract and vortexing at high speed for 2 minutes. Measurement was taken 24 hours later. The emulsification index(E24) was determined as the percentage of height of the liquid column. The E24 was measured in three replicates for each surfactant (Schulz et al., 1991). The surfactant content is correlated with E24. Evaluating emulsification capacity is a basic screening method that is appropriate for the initial screening of surfactant generating microorganisms (Plaza et al.,2005).
E24 = h emulsion × 100%
————
h total
Characterization of Surfactants
Using High performance liquid chromatography, surfactant samples were analyzed using HPLC system 1260LC syringe, filter 0.2cm:0.4mm equipped with proshell 120EC, C18 4 micrometer, column gravity= 150× 4.6mm particle size 3-micrometer, machinery-nongel was used with DAD (VWD) diode array detector 257 nanometer, data acquisition was performed using chemostation software (Agilent Cooperation). Flow rate was set up at 0.700ml/nm and the column oven temperature was set to 28ºC. The mobile phase solution; Acetonitrite A and 0.1% formic acid in ultra-pure water B were used and running buffer. The method started with a ratio A:B from 70-30% and a linear gradient was applied to reach a ratio of 80:20% in 8 mins. The acetonitrile fraction was increased linearly from 80-100% between 9 and 10 mins and decreased linearly to 70% in 11 and 12.5 mins. The injection volume of sample was set at 20vl detected through a DAD (VWD) detector at 257mm. Each analysis was completed within 15mins.
Results
The isolation of diesel degrading bacteria from the leaves of Celosia argentea, resulted in the isolation of two competent diesel degrading organisms (Table 1). The isolates were identified using biochemical and 16S rRNA molecular typing as Lysinibacillus sphaericus (strain Bibi1) and Proteus mirabilis (strain Bibi8)
Table 1: Biochemical characterization of isolates
| Isolate code | Gram staining | Catalase | Indole | Simmon citrate | motility | oxidase | Hydrogen sulphide | Putatitve identity |
| BiBi1 | +ve | +ve | -ve | +ve | +ve | +ve | +ve | Lysinibacillus sphaericus |
| Bibi8 | -ve | +ve | -ve | +ve | +ve | -ve | +ve | Proteus mirabilis |
The growth profile of the isolates on diesel as monitored via observation of physical turbidity and the disappearance of the diesel contaminant in the enrichment flask (Figure 1A and 1B). Strain BiBi1 grew exponentially from day 0 to Day 8 (OD = 0.7nm), whereas growth where at peak at Day 10 (OD = 0.8 nm). It was observed that a steady gradual decrease in the pH of the medium from 7.2 day 0 to 6.3 day 13. Also Proteus mirabilis had a steady growth rate on diesel from day 0 to day 10 whereas growth was at its peak on day 12 (OD = 0.85 nm), acidic pH was also observed in the growth media with a steady fall in pH from 7.2 (day 0) to 6.2 (day 13).

Figure 1A: Growth dynamics of Lysinibacillus sphaericus on diesel

Figure 1B: growth dynamics of Proteus mirabilis on diesel
The potential of the isolates to produce surface active biomolecules were tested via hemolytic and E24 test (Table 2). Strain BiBi1 showed incomplete zones of clearing on sheep blood agar which signify alpha hemolysis, whereas BiBi8 showed complete zone of clearing signifying beta hemolysis. Although hemolysis test is presumptive for biosurfactant production. Also, the emulsification index (E24) test showed that strain BiBi1 had E24 values of 51.28% whereas BiBi8 showed 76.32%. the values were remarkably high for a hydrocarbon utiliser since organism produced surface active molecule in response to degradation of hydrocarbon in the environment. It also implies that the organisms can be used to control the menace of hydrocarbon pollution in our local environment
Table 2: Biosurfactant producing capability of isolates
| Isolate code | Hemolytic assay | Emulsification indices (E24) |
| BiBi1 | Alpha hemolysis | 51.28 |
| BiBi8 | Beta hemolysis | 76.32 |
Following biosurfactant production, the nature of the surface active biomolecule produced by diesel degrading isolates were characterized using the high performance liquid chromatography (HPLC) (Figure 2A and 2B). Lysinibacillus sphaericus produced rhamnolipids and subtilisin as its surface biomolecule although rhamnolipid was produced more than the former. Also the production of rhamnolipid by Proteus mirabilis was massive (Figure 2B) as compared to with Lysinibacillus sphaericus, in addition to rhamnolipid, lipopeptides was also produced by strain BiBi8.
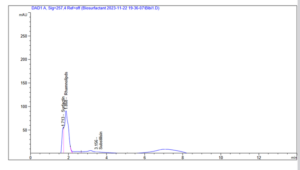
Figure 2A: HPLC profile of the Rhamnolipid and Subtilisin biomolecules produced by Lysinibacillus sphaericus
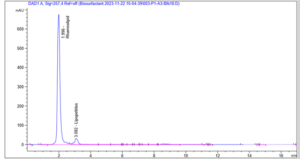
Figure 2B: HPLC profile of the rhamnolipid and lipopeptide biomolecules produced by Proteus mirabilis
Studies over the years have confirmed that bacteria present in aquatic and terrestrial environments can be excellent hydrocarbon degraders. In recent times, there has been research looking into bacteria present on the leaf surface and their potentials to degrade hydrocarbons and produce surfactants. Biodegradation is known as a major mecahnism for removing recalcitrant hyrocarbons in the environment. The organisms used in this study have been well studied with excellent results ( Hernandez-Santana and Dussan, 2018; Ke et al., 2021; Shettima et al., 2023). This study further proves that phyllosphere bacteria are hydrocarbon utilisers and surfactant producers. In the study two diesel degrading bacterial identified using biochemical and molecular typing were Lysinibacillus sphaericus strain BiBi1 and Proteus mirabilis strain BiBi8 (Table 1). Isolating the bacterial strain showing good degradation capacity on diesel is not surprising as findings from this study corroborated previous studies when tested for their ability to utilise diesel as source of carbon and energy and also produce surfactants which will enhance their bioavailability to the hydrocarbon (Oso et al., 2019; Oso et al., 2021). The bacterial strains were first grown on LB agar which does not determine their hydrocarbon utilising ability. This study therefore establishes that bacteria present on Celosia argentea can be excellent hydrocarbon degraders which can aid the removal of this recalcitrant from the environment. Many bacteria have been studied extensively in degradation studies and surfactant production, Pseudomonads however remains one of the most studied bacteria. As observed in Burch et al., 2014, the surfactant produced by Pseudomonas syringae B728a helped the organism thrive in the phyllosphere when the environment becomes unfavourable. This is achieved when the surfactant breaks the surface tension of water and lower the contact angle of water droplet on the leaf. This in turn increases the area where the water droplets cover on the leaf and allow for the movement of the bacteria and for nutrients to diffuse toward the bacteria. Surfactant production in the phyllosphere also increases wettability and permeability of leaf cuticle. This shows that surfactants on leaf surface are beneficial to epiphytic bacteria in the phyllosphere.
To test for surfactant production in Lysinibacillus sphaericus and Proteus mirabilis, two complementary assays were used i.e. drop collapse method and emulsification index (Ilori et al., 2005; Oso et al., 2019; Wu et al., 2022). Both methods indicated the presence of surfactants in the bacterial strains. Although, the drop collapse assay proved to be a faster detection method in comparison to emulsification index which supports studies from Burch et al., 2014 and Oso et al., 2019 had previously studied surfactant production in phyllosphere bacteria isolated from spinach and romaine lettuce (Fig 2A and 2B, Table 2). In terms of hydrocarbon utilisation, both strains showed to be hydrocarbon utilisers when grown in MSM supplemented with diesel while no growth was observed in the control flasks which contained no carbon source. This further confirms that these bacterial strains are hydrocarbon utilisers as carbon source and and surfactant producers based on previous studies which also show the presence of alkane degrading genes alkB in trees found in urban areas (Gandofi et al., 2017). The results obtained broadens the understanding of the presence of hydrocarbon utilisers in the phyllosphere and can contribute to the creation of an effective biodegradation programme, thereby contributing to a greener environment.
The phyllosphere has been proven to be an excellent habitat for hydrocarbon utilisers and surfactant producing bacterial strains. The strain used in this study were able to utilise bacteria and also produce surfactants. It is however unclear if these strains are able to utilise the long chain aliphatics present on the cuticular waxes. This will be looked into in the future. A thorough investigation into biodegradation studies will also be carried out to determine what components of diesel the bacterial strains able to degrade.